
Reich, D. (2018, March). How genetics is changing our understanding of race. The New York Times.
Neiman, A. (2010). Race: A social deconstruction of a biological concept. Biology and Philosophy, 25(1), 123–135.
Taboo genetics. (2013) Nature, 502(7470), 15–17.
Baker, J. R. (2012). Race (3rd ed.). Oxford University Press.
Mayr, E. (2002). The biology of race and the concept of equality.
Raymond, M. (2013, May 17). Sex and races: Two biological realities. Le Monde. (Original work published in French)
1. Introduction
2. Classification of Homo Sapiens in the Animal Kingdom
Table 1: Taxonomic Classification of Homo sapiens
3. The Main Human Races
Table 2: A Dozen of Human Races
4. The Emergence of the Main Human Populations/Races
-Africans
-North Africans and South Asians (Middle Easterners)
-Southeast Asians
-Pacific Islanders
-Australian Aborigines
-Europeans
-East Asians
-Arctic People
-Native Americans
5. Invalidation of the Main Counterarguments
6. The Genomic Revolution and Racial Ancestry
7. Medication and Genetic Variability across Races
8. Conclusion
“It is no longer possible to ignore genetic differences between races. (…) Some claim these differences are minor, but that is incorrect.” —David Reich, Harvard professor of genetics, 2018
“Those who claim there are no human races are evidently ignorant of modern biology. Races are not uniquely human; they exist in many animal species.” —Ernst Mayr, 2002 (one of the greatest evolutionary biologists of the 20th century)
“Claiming human races don’t exist as separate subspecies insults me and the objective work of evolutionary biologists. Modern progressives rejecting evidence contradicting their worldview is alarming. Society must reconsider such biased thinking.” —Richard Dawkins, 2005
“Educated, intelligent people may react harshly upon realizing the media and even academia have misled them about racial differences their entire lives.” —Steven Pinker, 2018
“It’s false that racial classification has no genetic or taxonomic significance. It’s false that two individuals from the same group differ as much as any two globally random individuals. It’s false that race cannot be predicted genetically.” —A.W.F. Edwards, Cambridge professor of genetics, 2003
In summary
It is a general principle of evolutionary biology that when populations of a species become isolated from each other, they evolve into two or more subspecies. They are called varieties, strains, or races. In the case of humans, these different varieties are called races. These different varieties evolve through four main processes: (1) founder effects, (2) genetic drift, (3) mutation, and (4) adaptation.
1. Introduction
The concept of human races and the associated notion of human equality have been subjects of considerable debate, particularly since the end of the Second World War. Over recent decades, the term “race” has been critically scrutinized and associated with strong negative connotations, often with the aim of discrediting its validity. Despite sustained efforts to remove the term from academic and public discourse, the underlying concept has persisted.
After a long period dominated by an egalitarian ideological stance, the idea of human races is once again widely acknowledged, although it is often expressed using alternative terminology such as “populations” or “genetic clusters” in contemporary scientific literature.
This historical example of ideological influence on scientific discourse can be seen as a clear case of Lysenkoism. As rational scientific inquiry into this topic re-emerges, it becomes increasingly important to re-examine past misconceptions critically.
2. Classification of Homo sapiens in the Animal Kingdom
The biological definition of a race or population:
A race or population is a subdivision of a species that inherits characteristics distinguishing it from other populations of the same species. In genetic terms, a race is defined as a population that differs in the frequency of certain genes compared to other populations—a consequence most often resulting from geographical isolation.
3. The Main Human Races/Populations/Genetic Clusters
It is not essential to refer to these genetic groups as “races.” While this term is the most common and scientifically established, alternatives such as “populations,” “genetic clusters,” or “subspecies” are generally considered synonymous and may be used interchangeably, depending on the context.
Table 2. Principal Races/Populations/Genetic Clusters/Subspecies in Homo sapiens: A Genetic Classification Based on a Sufficient Number of Polymorphisms.
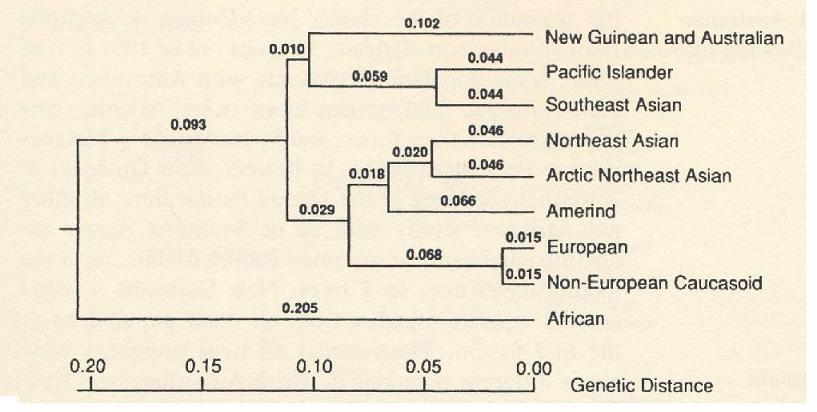
The number (n) of races, populations, or genetic clusters is variable. A race can be likened to a branch of a tree. How many branches does a tree have? If you observe the large branches near the trunk, you will see only a few major branches. As you move toward the periphery, you encounter increasingly smaller branches. This analogy aligns with the biological concept of race. Scientists typically consider the larger branches as representing races within a species. In the case of Homo sapiens, there are generally considered to be about a dozen such groups, although the exact number depends on the criteria used.
A subspecies or race (or genetic cluster or population) arises spontaneously through speciation processes, usually affecting a geographically isolated population. Prolonged isolation, coupled with specific environmental selection pressures and limited gene flow, leads to evolutionary changes—namely, modifications in allelic frequencies and, in some cases, the emergence of new mutations. These changes help align the phenotype with the surrounding environment through natural selection. When isolation is sustained over a long enough period, it may ultimately result in the formation of a new species. A race represents an intermediate stage in this evolutionary process. In fact, Darwinian evolution is not possible without the existence of races.
For a race to emerge, geographical isolation must persist long enough to allow for significant variations in allelic frequencies. The emergence of entirely new alleles is not strictly necessary—although such cases do exist. For example, alleles responsible for epicanthic folds are common in East Asian populations but absent in Europeans. Similarly, the presence of a distinct subcutaneous fat layer in East Asians gives their skin a slightly yellowish appearance. However, the existence of qualitative differences is not a prerequisite; sufficient quantitative variation, resulting from extended geographic isolation, is enough to define a race.
This raises the question: Have human groups remained sufficiently isolated from one another to justify the concept of distinct races? What is the minimum time t required for such differentiation to occur?
To approach this objectively, it is useful to examine how long it typically takes for races to emerge in various animal species, and then compare those durations to the periods during which human populations have remained isolated from one another.
For example, dogs diverged from wolves approximately 15,000 years ago and have since developed a remarkable diversity unmatched among mammals. Not only have their physical traits evolved, but their behavior has also changed significantly: dogs are far better than wolves at recognizing human voices and gestures, whereas wolves do not understand us at all.

There is no complex behavior observed in dogs that does not have a detectable precursor in wolves. Even if this is the case—and even if no entirely new mutations have been identified in the canine genome—this does not imply that all dogs are the same. Differences arise from variations in the frequencies of alleles within the gene pool. For example, an American study on dog attacks conducted between 1982 and 2006 reported approximately 1,100 attacks attributed to pit bull terriers, compared to only a single attack by a border collie.
Although all dogs belong to the same species, there are both morphological and behavioral (“psychological”) differences between breeds. These differences are also evident in learning ability: the number of repetitions required to learn a new command can vary significantly across breeds. A border collie, for instance, may learn a new command after just five repetitions and respond correctly 95% of the time, whereas a basset hound might require 80 to 100 repetitions and respond correctly only about 25% of the time.
Let us return to Homo sapiens and briefly trace human evolution to demonstrate that the durations of isolation between racial groups are comparable to those observed in other animal species. As illustrated in Figure 2, these periods of isolation are not uniform across all human groups. For example, African populations have been isolated from other human groups for approximately 100,000 years, whereas East Asians diverged from other populations over 40,000 years ago, making them genetically closer to certain other groups.
While the example of domestic dogs may be familiar, it is not necessarily the most appropriate for comparison, as it involves artificial selection imposed by humans. It serves merely as an illustrative case. In contrast, most animal species develop subspecies or races through natural speciation processes.
The giraffe, for instance, has nine recognized races or subspecies:
1 C. peralta – Southwest Niger.
2. c. reticulata – Northeastern Kenya, Ethiopia, Somalia.
3. c. Angolensis – Angola, Botswana, Namibia.
4. c. antiquorum – Chad, Central African Republic, Northeastern Cameroon.
5. c. tippelskirchi – Masai Giraffe – Central and Southern Kenya, Tanzania, Eastern Rwanda
6. c. camelopardalis – Eastern Sudan, northeastern Democratic Republic of Congo.
7. c. rothschildi – Uganda, North-Central Kenya.
8. c. giraffa – South Africa, Botswana, Zimbabwe, Mozambique.
9. thornicrofti – Zambia.
The wolf has 38 recognized subspecies or races.
The fox has 45 subspecies or races.
The raccoon has 22 subspecies or races.
The Bornean orangutan has 3 subspecies or races (in addition to several extinct ones).
Kangaroos have 2 subspecies or races occupying distinct territories within Australia.
In all these cases, the fundamental processes of speciation are at work. The emergence of a dozen races or subspecies within Homo sapiens does not deviate from these same evolutionary mechanisms.
4. The Emergence of the Main Human Races/Populations.
Homo sapiens emerged in Africa approximately 200,000 years ago.
1. Africans
Homo sapiens originated around 200,000 years ago in equatorial Africa.
2. North Africans and South Asians (Middle Easterners)
The first groups to migrate out of sub-Saharan Africa settled in North Africa and Southwest Asia between 100,000 and 90,000 years ago. These populations were geographically isolated from sub-Saharan Africans by both distance and the Sahara Desert, and they gradually evolved into a distinct racial group encompassing North Africans and South Asians.
3. Southeast Asians (Cambodia, Indonesia, etc.)
Around 70,000 years ago, people from South Asia migrated to Southeast Asia, where they evolved into the Southeast Asian population.
4. Pacific Islanders
Approximately 6,000 years ago, some Southeast Asians began migrating into the Pacific Islands. Due to prolonged geographic isolation, they developed into a distinct group: the Pacific Islanders.
5. Australian Aborigines
Groups from South and East Asia migrated through the Indonesian archipelago and reached New Guinea around 65,000 years ago. About 60,000 years ago, some of them reached Australia, where they evolved into the Aboriginal Australians. A closely related group remained in the highlands of New Guinea, known today as the Papuan Aborigines.
6. Europeans
Some populations who had previously colonized the Near East (between 100,000 and 90,000 years ago) migrated northward. Around 60,000 years ago, they reached the Caucasus and later spread into Ukraine, and about 40,000 years ago, into Central and Western Europe. Other groups from Southwest Asia began colonizing southeastern Europe through Anatolia. These populations gradually evolved into Europeans, characterized by lighter skin and, in northern regions, features such as blonde hair and blue eyes. They were isolated from South Asians and North Africans by the Mediterranean Sea, and from the east by the Black and Caspian Seas, the Caucasus Mountains, the Himalayas, and the Karakum Desert in present-day Turkmenistan.
7. East Asians (China, Japan, Korea, Singapore, etc.)
Populations from South and Central Asia began moving into Northern Asia between 60,000 and 50,000 years ago, where they evolved into East Asians. Geographical barriers such as the Gobi Desert and the Himalayas isolated them from Europeans and South Asians. These populations experienced colder winters—often reaching -12°C during glaciations—which led to adaptations such as flatter noses (to reduce frostbite risk), shorter limbs, thicker torsos, and subcutaneous fat layers that gave the skin a yellowish appearance (similar to Arctic peoples). Facial hair became less common among men due to frostbite risk, and the eye shape (epicanthic fold) evolved to reduce exposure to icy wind.
8. Arctic Peoples
Between 50,000 and 40,000 years ago, some groups migrated into the far north of Asia, where they evolved into Arctic populations. These groups became isolated from East Asians by mountain ranges such as the Chersky, Khingan, and Sayan, as well as vast forests and rivers. They experienced extremely cold winters—around -15°C, dropping to -20°C during the Würm glaciation—and developed characteristic cold-adapted traits: flat noses, short limbs, thick torsos, subcutaneous fat layers, and ocular adaptations.
9. Native Americans
Native Americans descended from populations that migrated from Northern Asia to Alaska via the Bering Strait. While traditionally believed to have crossed around 12,000–11,000 years ago, genetic and archaeological evidence suggests a much earlier migration—around 40,000 years ago. Artifacts dated to 24,000 years ago in Mexico, 30,000 years ago in California, and 33,000 years ago in Monte Verde, Chile, support this earlier timeline. These migrants gradually spread across the Americas, evolving into the Native American populations.
Genetic evidence confirms their shared ancestry with East Asians: both groups exhibit similar hair texture, shovel-shaped incisors, and skull morphology. They also share rare blood markers, such as the Diego antigen and low prevalence of Rh-negative blood types.
Conclusion: Isolation and Evolution of Races
The durations of isolation—approximately 100,000 years between Europeans and Africans, and 40,000 years between East Asians and Europeans—are sufficient to explain the development of distinct human races. These groups evolved in vastly different environments, with unique climates and selective pressures, leading to clear physical differences. The contrast is often so marked that individuals from one population are not easily confused with those from another—for instance, no European is mistaken for an African, or vice versa.
Although it is often claimed that differences between human populations are only superficial (e.g., skin or hair color), many studies show otherwise. Experts can often determine an individual’s ancestry from skeletal remains, indicating that differences extend to the bone structure. Genetic research has identified variations not only in outward appearance but also in traits like brain development and metabolic processes.
While biology naturally focuses on visible traits, this does not mean that invisible differences do not exist. In fact, superficial differences are often just the most apparent expression of deeper genetic variation.
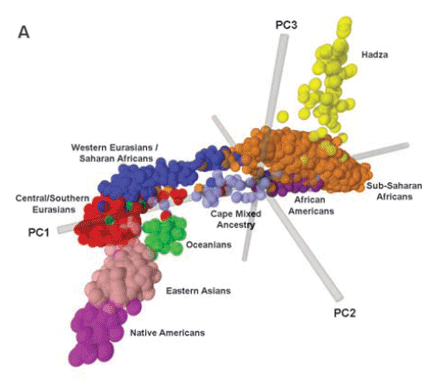
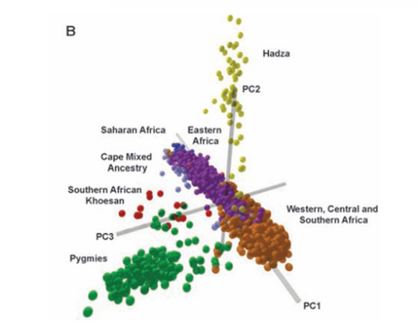
Within a race, there are also subsets. Take the example of the race of sub-Saharan Africans:
John Goodrum has compiled comparative data on genetic diversity in animals, including Homo sapiens. This diversity is assessed in terms of heterozygosity. The average autosomal microsatellite heterozygosity of a population (H) is the percentage of heterozygous individuals (two alleles) at a random locus. H varies from 0 to 1 or 0-100%; the higher the value, the greater the genetic diversity of a species. In the table below, we find the theoretically expected heterozygosity, He, and the heterozygosity observed in practice, Ho.
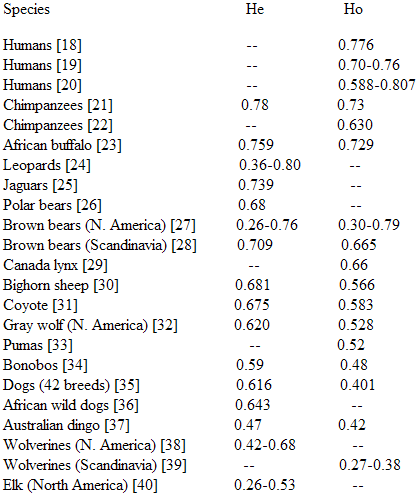
Homo sapiens is genetically more diverse than many mammal species.
Indeed, many species, including subspecies or races, have a lower value of diversity between these racial subpopulations. Goodrum has compiled examples of genetic diversity between races in different animals, the genetic distance is in FST. In the table below, note that 0.168 is the same as 16.8%, 0.155 is the same as 15.5%, etc.

Homo sapiens is therefore in a position comparable to many other animal species.
The following table, based on the Sewall Wright scale, shows again that Homo sapiens, with a value of 15.5, does not differ markedly from many other species in the animal kingdom.
| FST | Extent of differentiation between populations |
| 0 – 0.05 | small |
| 0.05 – 0.15 | moderate |
| 0.15 – 0.25 | great <-homo sapiens (>0.15) |
| > 0.25 | very great |
5. Invalidation of the Main Counter-Arguments
Let us now examine the most frequently cited counter-arguments against the concept of race, and offer responses rooted in genetics and biology.
1. “There are biological differences among human populations, but they are not sufficient to justify speaking of races.”
Response: Either the term “race” is applicable to all biological species, including Homo sapiens, or it must be abandoned entirely. As previously discussed, the estimated isolation times between Africans and Caucasians are around 100,000 years, and between East Asians and Caucasians, approximately 40,000 years. It is illogical to assume that the genetic variation resulting from this isolation would be less significant in Homo sapiens over 100,000 years than in dogs over 15,000 years.
Moreover, Homo sapiens and chimpanzees share 98.8% of their genetic material, yet no one would deny the substantial phenotypic and cognitive differences between them. Humans and Neanderthals share 99.5% similarity, and men and women even more. Still, these differences result in clearly recognizable distinctions.
Thus, even very small genetic differences can lead to significant phenotypic, behavioral, and physiological variation. The variability among human populations, due to sufficient isolation, is enough to justify discussions about race.
2. “We are 99.9% genetically identical.”
Response: This statement is technically correct, but misleading. The human genome contains roughly 3.2 billion base pairs, so a 0.1% difference still represents about 3.2 million individual genetic variations. These variations can have significant effects.
Humans share over 90% of their genes with mice, yet no one argues they are essentially the same. Men and women share over 99% of their DNA, yet sex is universally acknowledged as biologically meaningful.
In addition, copy number variations (CNVs)—large segments of DNA that may be duplicated or deleted—comprise about 12% of the human genome. Some CNVs involve millions of base pairs and affect thousands of genes. CNVs are increasingly associated with a variety of neurological and physical disorders, making the 99.9% claim overly simplistic and biologically outdated.
3. “Human variation is continuous, such as in skin color, and does not form discrete races.”
Response: While continuous variation exists due to interbreeding, this does not invalidate the existence of races. Just as blending red and yellow produces orange without eliminating the existence of red and yellow, intermediate forms support rather than contradict the existence of discrete genetic populations.
In reality, large-scale interbreeding is limited mostly to regions such as South America. In much of Asia, Africa, and Europe, population groups have remained relatively distinct. Rosenberg et al. (2005) showed that even with gradual changes, genetic clustering based on geographic ancestry remains clear.
4. “Racial classification is arbitrary and based on superficial criteria.”
Response: Racial classification is not arbitrary; it reflects patterns of genetic similarity resulting from geographic isolation. The genetic differences among groups have biological significance—such as higher melanin levels in Africans, variations in testosterone, bone density, gestational length, onset of puberty, and rates of early development.
Moreover, laboratories can determine ancestry from samples of blood, hair, bones, or other tissues. These markers are based on genetic traits that correlate with ancestral geographic regions.
5. “Race is often confused with species.”
Response: This is a common misunderstanding. All humans belong to the same species, Homo sapiens, but species can—and frequently do—contain subspecies or races. These are subdivisions based on historical separation and adaptation to local environments.
6. “Arguments from authority should be trusted over personal observations.”
Response: While expert consensus is valuable, critical thinking and open debate are essential to scientific progress. Disregarding observable and well-supported data on the basis of ideological preference is intellectually dishonest. Many scientists now prefer terms such as “ethnic group” or “genetic cluster,” but the underlying biological distinctions remain unchanged.
7. “The concept of race is obsolete and rooted in colonial history.”
Response: Science has frequently evolved past outdated or misused concepts without discarding entire fields. For example, the misuse of physics to develop nuclear weapons did not result in the abandonment of physics. Similarly, racial classification should not be dismissed due to past ideological misuse.
8. “Race is a political and social construct, not a biological reality.”
Response: While social perceptions of race are shaped by culture and politics, biological distinctions between human populations do exist. Ironically, the assertion that race is purely a social construct is itself an ideological narrative developed in recent decades.
9. “Censorship prevents open scientific debate on race.”
Response: Because arguments against the concept of race can often be refuted, public discourse is increasingly censored. Dissent from the mainstream narrative is discouraged, and ideological conformity has, in some circles, replaced objective inquiry.
10. “Denying the concept of race is necessary to fight racism.”
Response: This is a moral argument, not a scientific one. Suppressing facts for ideological reasons mirrors the logic of past censorship—such as denying heliocentrism for fear of societal disruption. As Goethe famously said, “I prefer a disturbing truth to a useful error.” Science must not bend to ideology, however well-intentioned.
11. “Genes may influence physical traits but not cognitive or psychological traits.”
Response: It is implausible that natural selection would affect every major physiological system—digestive, cardiovascular, immune—except the brain. Research has demonstrated ongoing human brain evolution (Meckel-Bobrov et al., 2005; Evans et al., 2005), and behavioral genetics has long shown heritability of cognitive traits and personality (Loehlin, 1992; Pennington, 2007).
12. “Lewontin’s argument disproves the biological basis for race.”
Response: Richard Lewontin (1974) claimed that 85% of human genetic variation exists within populations, not between them, suggesting race was insignificant. However, A.W.F. Edwards (2003) rebutted this in “Lewontin’s Fallacy,” showing that while individual loci may not be informative, combined analysis of multiple loci allows for extremely accurate classification. Luigi Luca Cavalli-Sforza used hundreds of genetic markers to identify major racial clusters. The power of such clustering comes not from any single gene but from patterns across many genes.
Final Note:
Once all scientific arguments have been presented and an opponent continues to reject them without rational engagement, the discussion often devolves into emotional responses—insults, accusations, or moral condemnations. At that point, it ceases to be a scientific debate and becomes a form of ideological bullying.
6. The Genomic Revolution and Racial Ancestry
Initiated in 1990 and completed in 2003, the Human Genome Project successfully sequenced the entire human genome. However, the reference genome initially produced was a composite derived from multiple anonymous donors. It was not until 2007 that the full genome of a single individual—Nobel laureate James Watson—was completely sequenced. Since then, the cost of genome sequencing has decreased exponentially.
The Human Genome Project cost nearly $3 billion and required 13 years of collaborative work by an international team. By 2025, it had become possible for private companies to sequence an individual’s entire genome for as little as $200 using nothing more than a saliva sample. The process is now fully automated and can be completed in just a few hours.
Other companies provide even more affordable services by focusing solely on sequencing variable regions of the genome, known as polymorphisms. By targeting only the most informative segments of DNA, these services can significantly reduce the cost.
The best-known company offering such services is 23andMe, a Google-affiliated firm, which provides ancestry and health reports for around $99.
23andMe’s analysis includes information on genetic predispositions to various diseases (such as Alzheimer’s, psoriasis, cancer, and obesity), physical traits (eye color, hair texture and color, birth weight, height), drug response (including antidepressants, beta blockers, statins), and—most relevant to this discussion—racial ancestry with a high degree of precision.
This capability provides a practical validation of the biological concept of race, defined as a “subdivision of a species that inherits characteristics distinguishing it from other populations.” In the genetic sense, a race is a population that differs in the frequency of certain genes from other populations, primarily due to geographic isolation over extended periods.
The genetic distinctions observed (see image below) would be impossible if races did not exist or if genetic traits were randomly distributed across all human populations. Instead, it is precisely the non-random distribution of genes that enables ancestry to be accurately determined from a genome.
Below is the 23andMe genetic ancestry report of Lilly Mendel, mother of the pioneering geneticist Gregor Mendel. Her ancestry is shown to be 99.9% European.
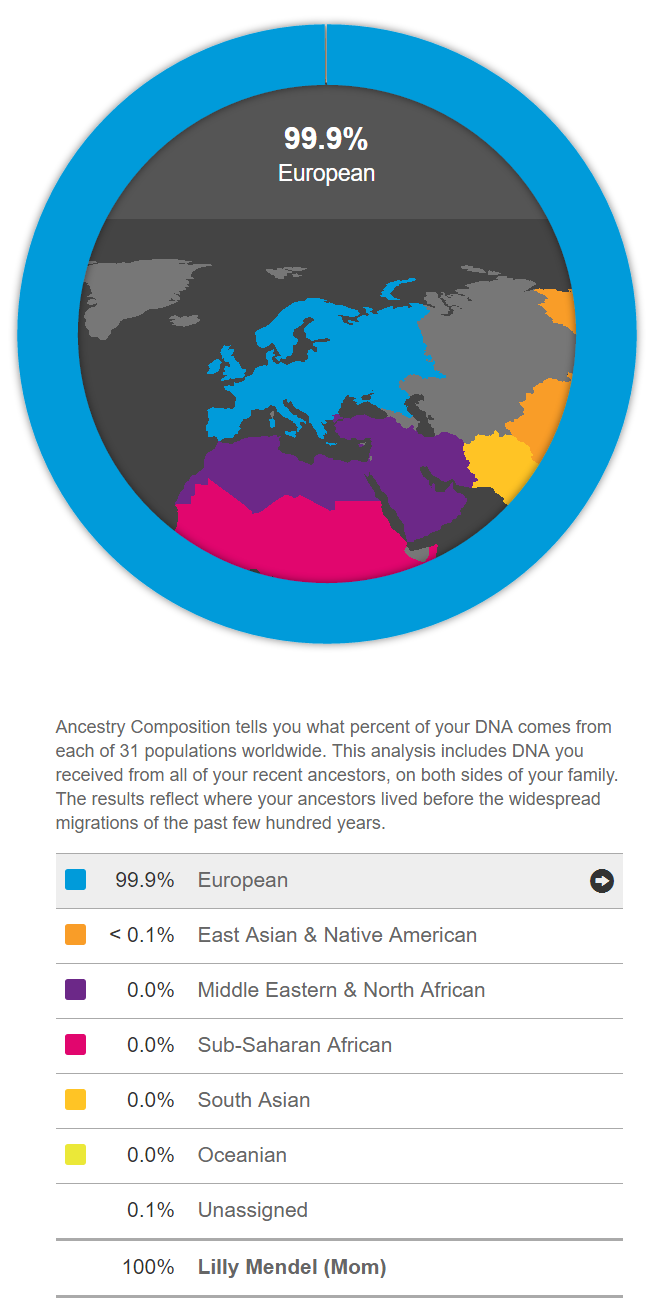
The initial analysis above provides a broad overview, identifying ancestry based on the major racial or genetic clusters. However, genetic analysis allows for much greater precision in determining ancestral origins.
Below is a more detailed report of Lilly Mendel’s genetic ancestry.
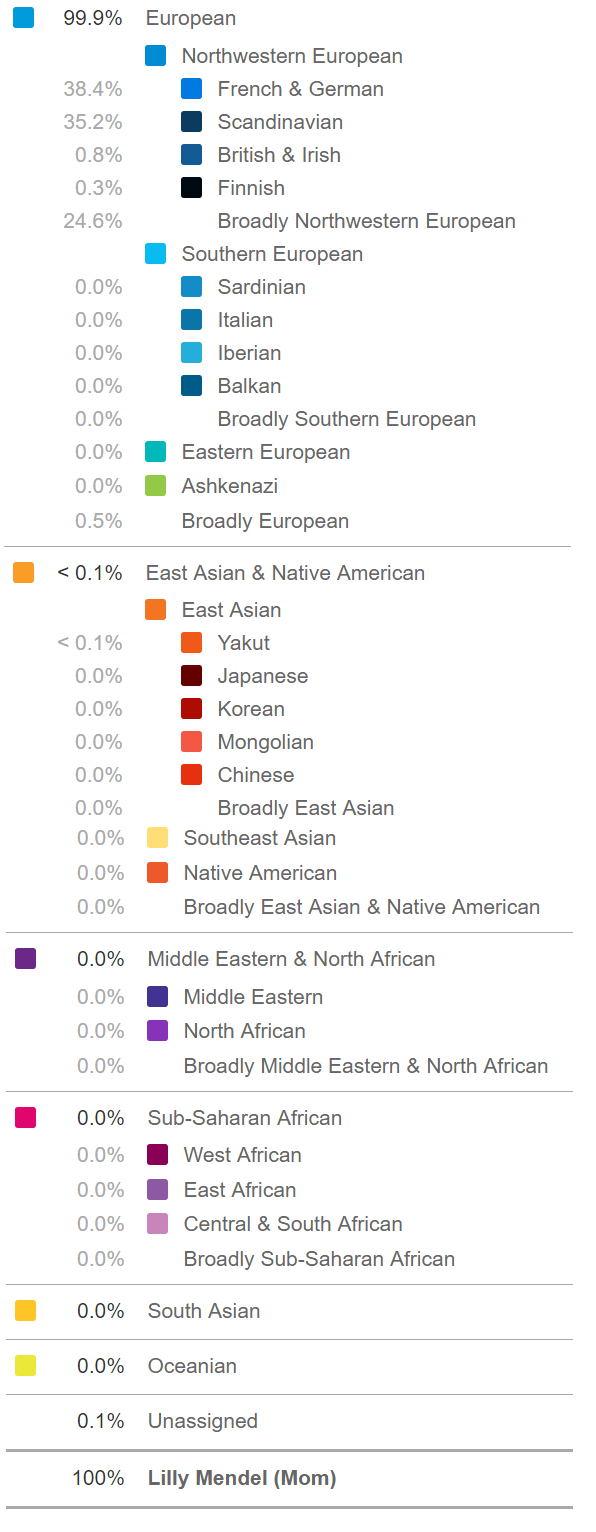
Such analysis would be impossible if racial or ancestral background had no genetic or biological basis, as some ideologues claim.
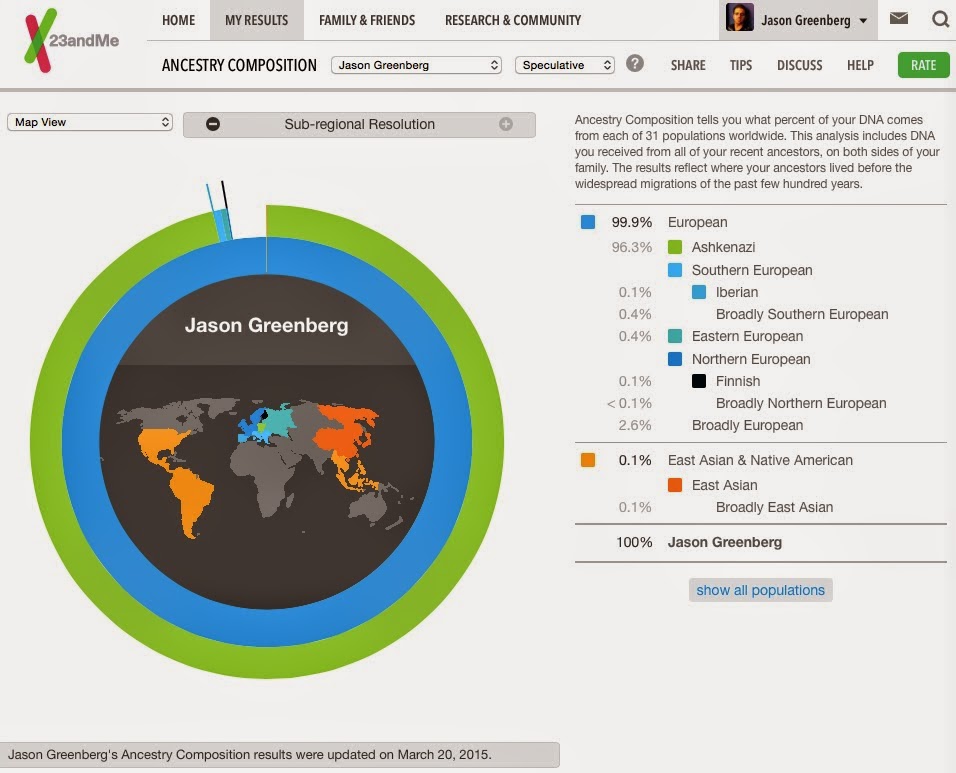
Below is a genetic analysis of an Ashkenazi Jew, a distinct subgroup within the European population. Ashkenazi Jews are notable for their high average IQ, estimated at around 110. Despite comprising a population of only about 10 million—the approximate population of Belgium—they account for approximately 50% of world chess champions, 50% of bridge champions, and nearly one-third of Nobel Prize laureates.
This elevated average intelligence is widely believed to have a hereditary component. Genomic studies consistently show that Ashkenazi Jews form a distinct ethnic group, with identifiable genetic patterns that set them apart from other European populations.
7. Medication and Genetic Variability Across Races
A Drug Specifically for African Americans: BiDil
The approval of BiDil was a watershed moment. On June 23, the U.S. Food and Drug Administration (FDA) authorized the release of the first medication marketed exclusively for African American patients. BiDil (short for “bi-dilators”) was found to significantly reduce mortality in African American patients suffering from advanced heart failure, according to a study published in The New England Journal of Medicine. This decision sparked intense debate about the emergence of race-based medicine.
Why does this happen?
The explanation lies in the genetic basis of drug metabolism. The efficacy of a drug is often closely tied to genetic factors, including allelic frequencies that may vary among racial groups. In the case of BiDil, a specific allelic distribution found predominantly in African Americans enhances the drug’s effectiveness.
This is not an isolated case. Numerous studies have demonstrated that the efficacy of certain medications—such as antihypertensives and antidepressants—can differ significantly between racial groups. For example, such medications often show reduced efficacy in individuals of African descent, while exhibiting increased efficacy in East Asian populations.
The Irony of Positive Discrimination
Some educational institutions apply race-based admission policies, and certain laws are designed to combat racial discrimination while simultaneously implementing forms of “positive discrimination” (affirmative action), often favoring underrepresented minorities or immigrants. It appears that the concept of race is invoked selectively, according to what aligns with social or political interests.
The Rise of Genetic Tools to Trace Ancestry
The technologies developed through the Human Genome Project are now widely applied to a variety of purposes. For example:
-
Companies offer African Americans the possibility of tracing their ancestral tribes (e.g., www.africanancestry.com).
-
DNA from bones discovered on a Russian farm confirmed a genetic connection to the Romanov family.
-
Genetic analysis has identified Thomas Jefferson as the likely father of one of Sally Hemings’ children.
-
The National Geographic Society’s Genographic Project allows individuals to trace their maternal or paternal lineage and understand their connection to prehistoric human migrations (www3.nationalgeographic.com/genographic).
Conclusion
The rejection of the biological validity of racial classification in Homo sapiens is largely ideological in nature. It is arguably a modern form of Lysenkoism—the Soviet-era practice in which science was distorted to serve political ends. Trofim Lysenko, a state-sponsored biologist, was notorious for suppressing genetics in favor of politically motivated theories on the malleability of human nature.
As the geneticist A.W.F. Edwards of Cambridge famously remarked in 2005:
“It is not true that racial classification has no genetic or taxonomic significance, or that one cannot infer a person’s race from their genes.” (Nature Genetics, p. 800)
A 2013 article in Nature titled “Ethics: Taboo Genetics” underscores this issue. It notes that research into racial differences in intelligence and personality has become a scientific taboo, driven more by ideological concerns than by scientific ones.
In a widely discussed New York Times article published in March 2018, Harvard geneticist David Reich also addressed this topic. He argued that acknowledging genetic differences between populations should not be equated with racism and that honest scientific discourse requires confronting these issues openly and responsibly.
“One may be concerned about possible misuse of data to justify racism, but as a geneticist, I also know that it is simply no longer possible to ignore the average genetic differences between races.
Revolutionary advances in DNA sequencing technology have been made over the past two decades. This progress allows us to measure with perfect precision what fraction of the genetic ancestry of an individual goes back, for example, to West Africa. With the help of these tools, we learn that, while race can be a social construct, the differences in genetic ancestry that are correlated with many current racial constructions are very real.
Well-intentioned people who deny the possibility of substantial biological differences between human populations curl up in an indefensible position, which will not survive the onslaught of science.
While most people agree that it is important to find genetic explanations for certain diseases, they are restive when it comes to genetic influences on behavior and cognition.
Is IQ, intelligence, or many years of education influenced by education? Of course. But does it also measure cognitive and behavioral characteristics? Almost certainly. And since all characters influenced by genetics differ from one population to another (since the frequencies of genetic variation are rarely identical from one population to another), genetic influences on behavior and cognition will also vary. one population to another.
You will sometimes hear that biological differences between populations are small because humans have diverged too recently from common ancestors so that substantial differences have emerged under the pressure of natural selection. This is not true. The ancestors of East Asians, Europeans, Africans, and Australians were, until recently, almost completely isolated from each other for 40,000 years or more, which is more than enough for the forces of evolution to work.
In the years to come, genetic studies will show that many traits are influenced by genetic variation and that these traits differ on average between human populations. It will be impossible – indeed, anti-scientific, silly and absurd – to deny these differences.”
-David Reich, Harvard University, March 2018.
References
Reich, D. (2018, March). How genetics is changing our understanding of race. The New York Times.
Neiman, A. (2010). Race: A social deconstruction of a biological concept. Biology and Philosophy, 25(1), 123–135.
Taboo genetics. (2013) Nature, 502(7470), 15–17.
Baker, J. R. (2012). Race (3rd ed.). Oxford University Press.
Mayr, E. (2002). The biology of race and the concept of equality.
Raymond, M. (2013, May 17). Sex and races: Two biological realities. Le Monde. (Original work published in French)